Competencia
La competencia es la interacción entre individuos de la misma especie (competencia intraespecífica) o de especies diferentes (competencia interespecífica) que utilizan el mismo recurso; éste suele estar en cantidad limitada. Como resultado de la competencia, el éxito biológico –o sea, el éxito en la reproducción de los individuos que interactúan puede verse reducido. Entre los muchos recursos por los cuales los organismos pueden competir se encuentran el alimento, el agua, la luz, el espacio vital, los sitios de nidificación o las madrigueras. La competencia puede ser por interferencia o por explotación.
Durante muchos años, la competencia ha sido invocada como una fuerza primordial en la determinación de la composición y estructura de las comunidades –o sea, el número y tipo de especies presentes y su distribución espacial y temporal dentro de la comunidad–. Sin embargo, recientemente diversos ecólogos han cuestionado la importancia de la competencia sobre la composición y la estructura de la comunidad. El debate –a veces cáustico– que se generó, concierne no sólo al papel de la competencia sino a los métodos que deben usarse para someter a prueba las hipótesis ecológicas.
Los investigadores que echaron las bases de la ecología moderna fueron atentos observadores de la naturaleza. Muchos de estos ecólogos creían, como tantos de sus continuadores, que la observación cuidadosa de los patrones naturales de distribución y abundancia de los organismos puede –sin manipulación experimental– producir los datos requeridos para someter a prueba las hipótesis sobre el papel que desempeñan la competencia y otras fuerzas para conformar la estructura de la comunidad. Para estos ecólogos, los patrones son los que hablan por sí mismos.
Otros ecólogos sostienen que los análisis experimentales en los cuales sólo se manipula una variable por vez son los únicos medios confiables para identificar los procesos que están ocurriendo realmente en la naturaleza.
En 1.934, el biólogo ruso Georgii Frantsevich Gause formuló lo que se conoció como el principio de la exclusión competitiva.
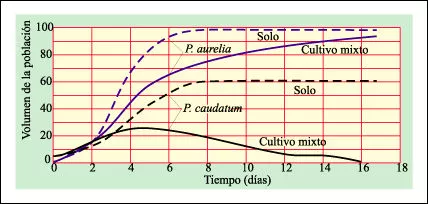
Experimentos de Gause con dos especies de Paramecium
Los resultados de los experimentos de Gause con dos especies de Paramecium demostraron el principio de que si dos especies se encuentran en competencia directa por el mismo recurso limitado –en este caso alimento– una elimina a la otra. Paramecium caudatum y Paramecium aurelia fueron cultivados primero separadamente en condiciones controladas y con un constante suministro de alimento. Como puede verse, P. aurelia creció mucho más rápidamente que P. caudatum, indicando que P. aurelia utiliza los suministros de alimentos disponibles de un modo más eficiente. Cuando los dos protistas fueron cultivados juntos, la especie que crecía más rápidamente superó en números y eliminó a la especie que crecía más lentamente.
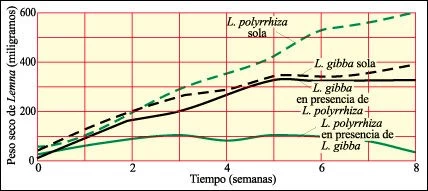
Un experimento con dos especies de lentejas de agua, diminutas angiospermas que se encuentran en estanques y lagos.
Una especie, Lemna polyrrhiza crece más rápidamente en un cultivo puro que la otra especie, Lemna gibba. Cuando son cultivadas juntas, Lemna gibba, que tiene diminutos sacos aéreos que la hacen flotar en la superficie, cubre a la otra especie y triunfa en la competencia por la luz.
El principio de exclusión competitiva de Gause nos llevaría a pensar que sólo podrían hallarse especies disímiles coexistiendo en las comunidades naturales. Pero, en realidad, es frecuente encontrar especies ecológicamente similares que viven juntas en la misma comunidad. Esta observación planteó la cuestión del grado de similitud que debe existir entre dos o más especies para que puedan continuar coexistiendo en el mismo lugar y al mismo tiempo. Esto llevó, a su vez, al concepto de nicho ecológico. Los análisis de situaciones en las cuales coexisten especies similares han demostrado que los recursos frecuentemente están subdivididos, o son distribuidos, por las especies coexistentes.
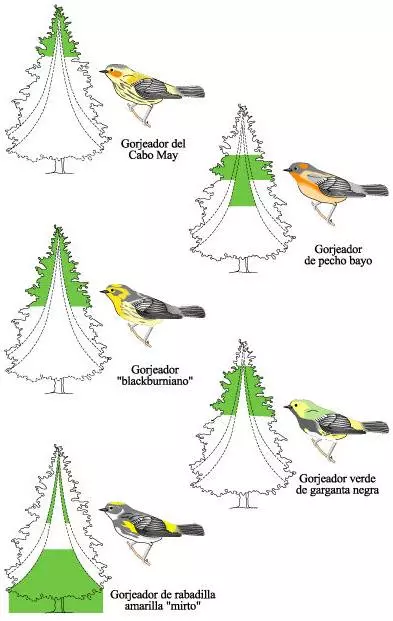
Las zonas de alimentación de cinco especies de gorjeadores norteamericanos en un abeto.
Las áreas coloreadas en el árbol indican en qué lugar cada especie pasa, al menos, la mitad de su tiempo de alimentación. Esta distribución de recursos permite que las cinco especies se alimenten en los mismos árboles.
El solapamiento de nichos describe la situación en la cual los miembros de más de una especie utilizan el mismo recurso escaso. En las comunidades en las cuales ocurre solapamiento en nichos, la selección natural puede dar como resultado un aumento de las diferencias entre las especies que compiten, un fenómeno conocido como desplazamiento de caracteres.
Uno de los ejemplos de desplazamiento de caracteres citado con más frecuencia es el del pico de los pinzones de Darwin.
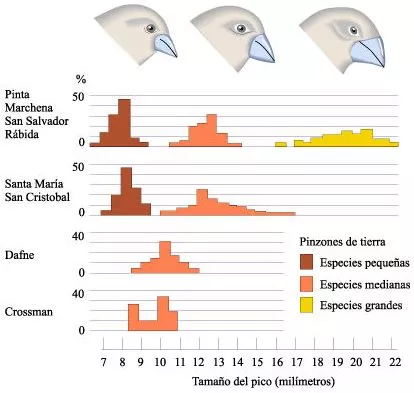
Tamaños de los picos en tres especies de pinzones terrestres de las Islas Galápagos.
Las medidas de los picos se muestran horizontalmente y el porcentaje de especímenes de cada especie se muestra verticalmente. Dafne y Crossman, que son islas muy pequeñas, tienen sólo una especie de pinzones terrestres cada una. Estas especies tienen tamaños de picos intermedios entre los pinzones medianos y los pinzones pequeños de las islas más grandes.
Los ecólogos han interpretado estos datos de dos maneras diferentes. Algunos sostienen que las diferencias observadas en el tamaño del pico son resultado de las presiones selectivas ejercidas por la competencia interespecífica. De acuerdo con ésta interpretación, la competencia entre los organismos cuyos nichos ecológicos se solapan causa una selección contra los individuos cuyas características se superponen, llevando a la divergencia observada entre las especies.
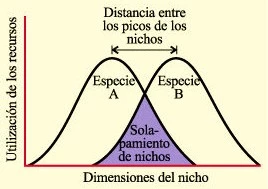
Gráfico de dimensión de un nicho ecológico
Las dos curvas con forma de campana representan la utilización de recursos por dos especies en una comunidad. La dimensión del nicho podría representar espacio vital como en el caso del musgo Sphagnum; espacio de alimentación, como en el caso de los gorjeadores; tamaño de las semillas comidas, como en el caso de los pinzones de las Galápagos, etc. La competencia es potencialmente más intensa en las áreas de solapamiento de nichos que lleva a la restricción de una o ambas especies en el espacio vital, el espacio para alimentación o el tamaño de las semillas ingeridas, etc. Se formula la hipótesis de que ésta competencia da como resultado una selección contra los individuos con características que se solapan llevando a la divergencia en los nichos de las dos especies.
Otros ecólogos, en cambio, señalan que es imposible determinar si los distintos tamaños de pico son el resultado de interacciones competitivas que ocurrieron en épocas en que las diferentes especies coexistían en las mismas islas, o si son el resultado de adaptaciones a las condiciones locales de épocas en que las especies se encontraban aisladas unas de otras en islas diferentes. Algunos ecólogos creen que la única evidencia indiscutible del desplazamiento de caracteres consistiría en demostrar que las especies en cuestión están divergiendo actualmente en las áreas donde se las encuentra juntas. Ambos grupos de ecólogos coinciden, sin embargo, en que cualquiera sea la causa evolutiva, las diferencias en el tamaño y la configuración del pico capacitan a las diferentes especies de pinzones para explotar diferentes fuentes de alimento y por consiguiente, para coexistir.
Prácticamente, todos los ecólogos coinciden en que la competencia ocurre realmente en la naturaleza con una intensidad que varía de acuerdo con la especie implicada, el tamaño de las poblaciones que interactúan y la abundancia o escasez de los recursos. Aunque el análisis de los patrones de distribución espacial y de recursos pueden dar idea de la existencia y la importancia de la competencia en una situación determinada, se necesitan experimentos que produzcan cambios en la composición de la comunidad para demostrar que realmente hay competencia.
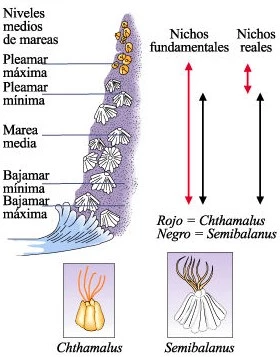
Competencia interespecífica entre Semibalanus y Chthamalus.
Las larvas de ambas especies se establecen en una amplia área pero los adultos viven en áreas precisamente restringidas. Los límites superiores del área de Semibalanus están determinados por factores físicos tales como la desecación. Los percebes Chthamalus, sin embargo, no viven en el área de Semibalanus, no por razones físicas (probablemente porque el área físicamente es menos limitante), sino por causa de los percebes Semibalanus. El Semibalanus crece más rápido y dondequiera que se encuentre con Chthamalus dentro de su propia área, o lo despega de las rocas, o crece por encima de él. Este estudio y otros análogos han generado los conceptos de nicho fundamental y nicho real. El nicho fundamental describe los límites fisiológicos de tolerancia del organismo; es el nicho ocupado por un organismo en ausencia de interacciones con otros organismos. El nicho real es aquella porción del nicho fundamental realmente ocupada; está determinada no sólo por los factores físicos sino también por las interacciones con otros organismos.
La mayoría de los estudios de competencia han puesto de relieve las adaptaciones y distribuciones de recursos que posibilitan la coexistencia de especies similares dentro de una comunidad. Sin embargo, ésta es una opinión sesgada, dado que es difícil estudiar las interacciones entre las especies después que uno de los protagonistas ha emigrado. Así como la competencia dentro de las especies lleva a la eliminación de la gran mayoría de los individuos, la competencia entre las especies puede llevar a la eliminación de una especie de la comunidad.
Bibliografía:
- Apuntes de clase - Facultad de Agronomía - U.B.A
- Helena Curtis - N. Sue Barnes. "Biología". Sexta edición en español
Autor: Diana Victoria Netto. Ingeniera Agrónoma. Argentina.
Editor: Ricardo Santiago Netto (Administrador de Fisicanet).
Competencia intraespecífica y competencia interespecífica.